


Les polymères biologiques microbiaux prometteurs : sources, arrière-plan génétique et applications techniques.
Les polymères biologiques
Les polymères biologiques, qui sont devenus précieux pour une utilisation dans de nombreuses applications, surpassent les dérivés pétrochimiques en étant biodégradables et compatibles sur le plan biologique et environnemental.
Les sucres sont un autre type de macromolécules largement présentes dans la nature. Ils peuvent être séparés en fonction de la morphologie de leur localisation en tant que partie de la membrane cytoplasmique ou sous forme de polysaccharides à l'intérieur des cellules dans la paroi cellulaire ou sous forme de polysaccharides extracellulaires en dehors de la paroi cellulaire.
Les sucres extracellulaires ont deux structures : ils ne s'attachent pas aux cellules, forment un mucus lâche et les rendent visqueux ; lorsque les bactéries se développent dans des milieux solides ou liquides, elles prennent une forme visqueuse et forment des capsules adhérentes à la paroi cellulaire ou des capsules microscopiques. Elles ont des structures et des limites uniques et ne se séparent que progressivement dans des solutions salines ou aqueuses. Par conséquent, les capsules microscopiques et les capsules peuvent être séparées du mucus lâche par centrifugation (Wilkinson, 1958).
Les polysaccharides extracellulaires sont des polysaccharides à longues chaînes composés d'unités de sucre ramifiées répétées ou de dérivés de sucre, principalement le galactose, le ramnose et le glucose en proportions variables. Ils sont divisés en deux groupes : les polysaccharides hétérogènes (gomme xanthane et gomme gélane) et les sucres homogènes (kurdlan, dévoraman, cellulose, glucane et amylopectine). (Sutherland, 1994)
Les polysaccharides homogènes sont constitués d'un seul type de monosaccharides (D-fructose ou D-glucose) liés par un nombre fixe de groupes de types de liaisons ou de types de liaisons individuelles. Les polysaccharides hétérogènes sont composés de plusieurs copies de sucres oligosaccharidiques contenant de 3 à 8 résidus produits par un autre organisme microscopique. Les sucres extracellulaires ont de nombreuses applications industrielles dans les industries pharmaceutiques et alimentaires ainsi que dans d'autres industries telles que les cosmétiques, les agents de cristallisation, les médicaments pour le traitement des plaies, le papier et les textiles (Sutherland, 1998).
Les exemples importants de monosaccharides sont la cellulose. La cellulose est le polymère biologique le plus important, et elle est principalement produite par les plantes. Cependant, certaines espèces de bactéries peuvent également produire de la cellulose avec une composition chimique similaire à celle de la cellulose végétale mais avec des propriétés physiques uniques. Cette différence est due au fait que le réseau fibrillaire de ses fibres microscopiques d'un diamètre de (0,1 µm) équivaut à environ 100 fois celle des fibres de bois.
De plus, contrairement à la cellulose végétale, la cellulose microbienne ne nécessite pas de transformation supplémentaire, étant débarrassée des contaminants et impuretés indésirables tels que la pectine, l'hémicellulose et la lignine, et de cette manière, elle a la possibilité de maintenir un niveau de polymérisation élevé (Nishi et al. 1990).
La cellulose microbienne
La cellulose microbienne a également montré des propriétés uniques, notamment en termes de pureté, de résistance à la traction, et un module de Young supérieur à celui des fibres synthétiques d'environ 30 à 40 % (Tanskul et al. 2013), ainsi que la capacité de gonfler jusqu'à 700 fois son poids sec. La structure de ses fibres est caractérisée par une douceur variant de 0,1 à 10 micromètres et une haute cristallinité, ainsi qu'une meilleure dégradation que la cellulose des plantes (Cai et Kim, 2010). En même temps, la cellulose extraite des plantes contient de la lignine, de la pectine et de l'hémicellulose comme composés principaux. Ce qui la rend plus adaptée comme matière première pour la production de papier de haute qualité, de confiseries et de haut-parleurs haute fidélité (Shoda et Sugano, 2005). Les fibres de cellulose bactérienne sont environ 100 fois plus fines que les fibres de cellulose végétale, ce qui en fait un matériau très poreux, permettant le transport de médicaments ou d'antibiotiques divers vers la lésion tout en agissant en même temps comme une barrière physique contre toute contamination externe. Par conséquent, elle est largement utilisée dans la guérison des blessures (Czaja et al. 2006).
La cellulose bactérienne est récemment devenue une caractéristique distinctive de nombreux produits biomédicaux importants, y compris les greffes vasculaires (tubes) ; les pansements pour les soins des plaies ; les fils de suture cultivés compatibles sur le plan biologique. Et les matrices de renforcement des tissus, ainsi que d'autres industries traditionnelles telles que le papier ; les produits alimentaires ; l'électronique et les produits cosmétiques. Il a été constaté qu'elle se distingue de la cellulose végétale en termes de régénération et de biodégradabilité, ainsi que de ses propriétés mécaniques uniques, y compris un degré de cristallinité élevé ; pureté ; capacité de rétention d'eau ; compatibilité biologique et flexibilité. La Colombie-Britannique est devenue une cible attrayante pour la production commerciale ces dernières années. (Naomi et al., 2020 ; Zhong C (2020)
Les souches bactériennes productrices de cellulose
Les organismes microscopiques qui produisent de la cellulose ont également des applications et des propriétés différentes de celles de la cellulose végétale. La cellulose est produite par divers organismes microscopiques tels que les algues, les bactéries et les champignons. Dans les algues vertes, le mannan, le xylane et la cellulose agissent comme structure de la paroi cellulaire des sucres. Bien que la cellulose soit produite en petites quantités dans la plupart des algues rouges (Rhodophyta), dans toutes les algues brunes (Phaeophyta) et finalement des algues dorées (Chrysophyta) (Richmond, 1991). Dans les Oomycètes, la chitine est complètement remplacée par la cellulose, représentant environ 15 % de la masse sèche de la paroi. Des espèces Gram-négatives telles qu'Alcaligenes, Azotobacter, Acetobacter, Sarcina, Rhizobium, Pseudomonas, Salmonella, Agrobacterium, Achromobacter, Aerobacter ont également été rapportées comme productrices de cellulose (Morgan et al. 2013).
De plus, la cellulose est synthétisée par la bactérie Sarcina ventriculi, qui est une bactérie Gram-positive, représentant environ 15 % de la masse totale des cellules sèches.
Synthèse de la cellulose bactérienne :
La synthèse de la cellulose bactérienne est un processus particulièrement précis et organisé qui implique de nombreuses étapes. Comme indiqué dans la figure 5 ; la structure supramoléculaire du complexe contenant un grand nombre de protéines catalytiques et régulatrices ainsi que des enzymes individuelles n'est pas encore claire. Les mécanismes et voies de synthèse du glucose-1,6-bisphosphate (UDPGlc) sont relativement bien connus, tandis que le mécanisme par lequel le glucose polymérise en longues chaînes non ramifiées doit encore être étudié (Prashant et al. 2009).
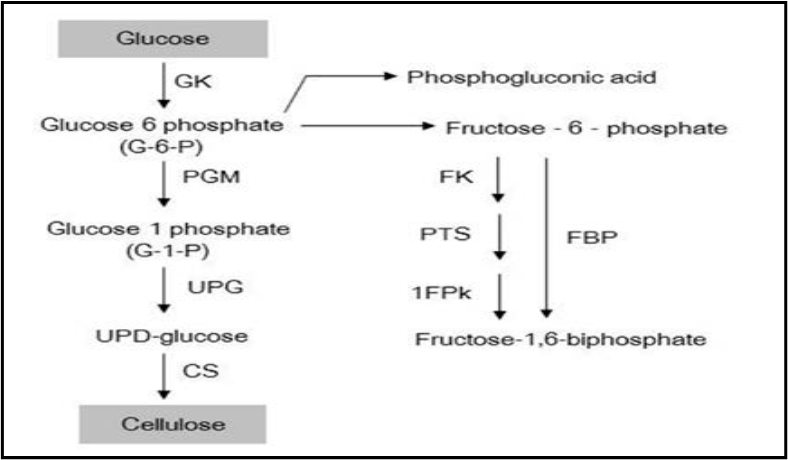
Figure : - Voie de biosynthèse de la cellulose bactérienne pour la bactérie Acetobacter sp. 1FPk (fructose-1-kinase), PTS (système de transport de phosphate), FBP (fructose-1,6-bisphosphate), FK (fructokinase)
Structure de la cellulose bactérienne
La formule moléculaire (C6H10O5)n de la cellulose bactérienne est la même que celle de la cellulose végétale, mais ses propriétés physiques et chimiques sont différentes. La structure de base de la cellulose bactérienne est un polymère de chaînes d'hydrogène étroitement liées entre les groupes hydroxyles, appelées microfibrilles, d'une épaisseur de 3 à 8 nanomètres et d'une largeur de 50 à 80 nanomètres, avec des chaînes de glucane connectées de l'intérieur vers l'extérieur. Cette structure fibrillaire fine de la cellulose bactérienne a été montrée pour la première fois en 1949 par Molithaler, qui a prouvé que chaque souche bactérienne produit une structure cellulosique différente.
Les gènes responsables de la production de cellulose microbienne
Dans les souches bactériennes productrices de cellulose. Notamment Gluconoacetobacter hansenii, le processus de biosynthèse est régulé par quatre gènes ; A, B, C et D qui comprennent l'opéron synthase de cellulose bactérienne (bcs) d'une longueur de 9,2 kilobases, précédé de fragments "régulateurs" qui sont dits nécessaires à la biosynthèse ; le gène cmcax qui code pour l'enzyme endo-β-1,4-glucanase qui dégrade BC et améliore sa synthèse, et le gène ccpAx qui est considéré comme jouant un rôle important dans le transport extracellulaire de (Thakur et al., 2020). L'enzyme synthase principale (PF03552) est codée par la partie bcsA, tandis que bcsB code pour une protéine régulatrice appartenant à la super-famille BcsB (pfam03170) qui code pour la protéine régulatrice de la biosynthèse de la cellulose et qui est considérée comme reliant le récepteur positif du di-GMP cyclique. (Yin et al., 2009) ; les deux comprennent une unité de glycosyltransférase complexe responsable du transfert de résidus de glucosyle du glucose UDP à la chaîne 1,4-glucane (Knott et al., 2016). En même temps, les parties bcsC (InterPro : IPR001440) et D (InterPro : IPR022798) sont considérées comme impliquées dans l'exportation et l'emballage des fibres de cellulose extracellulaire en formant un canal vers la membrane externe permettant la cristallisation. (Mehta et al., 2015). De nombreuses études ont rapporté que les fragments bcsAB seuls sont suffisants pour produire efficacement in vitro (Omadjela et al., 2013) ; cependant, d'autres études ont confirmé la nécessité des quatre gènes pour atteindre une production maximale in vivo (Vijayendran et al., 2020).
De nombreuses études ont examiné la production à grande échelle de BC dans de nombreux systèmes basés sur des plasmides (Omadjela et al., 2013 ; Costa et al., 2017 ; Buldum et al., 2018). Dans ce domaine, la plateforme E. coli est largement utilisée comme producteur de protéines de haute qualité en raison de sa cinétique de croissance rapide ; manipulation génétique simple et expression rapide des protéines, etc. (Sezonov et al., 2007). Les hôtes bactériens les plus couramment utilisés pour la production de protéines recombinantes à grande échelle sont les souches E. coli B [Rosetta et BL21 (DE3)] et les souches K-12, y compris la souche de réplication de plasmide principale DH5α (Marisch et al., 2013). Cependant, en tant que protéine liée à la membrane ; il a été rapporté que la surexpression de la synthase de cellulose dans E. coli BL21 (DE3) est toxique, ouvrant la voie à d'autres hôtes alternatifs tels que C41 (DE3). Une souche mutante dérivée de BL21 (DE3) a été utilisée pour étudier la possibilité d'une expression massive de protéines membranaires toxiques de l'opéron bcs. (Buldum et al., 2018)
Résumé
La cellulose bactérienne est un matériau biologique respectueux de l'environnement et biodégradable, avec des propriétés uniques qui lui ont valu une attention croissante pour de nombreuses applications industrielles. Cependant, la production à grande échelle de cellulose bactérienne en utilisant le type sauvage producteur de cellulose fait encore face à des défis en raison de la lenteur de la cinétique de croissance, de la grande quantité de glucose requise pour une production efficace, de la forte propension aux mutations spontanées associées aux fluctuations des conditions de culture/nutriments qui la rendent non productive. mutations, et une faible productivité de cellulose.